
Effect of extremely low-frequency magnetic fields on light-induced electric reactions in wheat - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC9176247/
Mar 5, 2025
10

Plant hormone transporters: what we know and what we would like to know - BMC Biology
link.springer.com/article/10.1186/s12915-017-0443-x
Mar 5, 2025
3

Phytohormone biosynthesis and signaling pathways of mosses - Plant Molecular Biology
link.springer.com/article/10.1007/s11103-021-01172-6
Mar 5, 2025
6

Day Length and Temperature Strongly Influence Sexual Reproduction and Expression of a Novel MADS‐Box Gene in the Moss Physcomitrella patens
onlinelibrary-wiley-com.ezpv7-web-p-u01.wpi.edu/doi/full/10.1055/s-2002-35440
Mar 5, 2025
1

Regulatory Mechanism Controlling Stomatal Behavior Conserved across 400 Million Years of Land Plant Evolution
www.cell.com/current-biology/fulltext/S0960-9822(11)00474-X
Mar 5, 2025
3
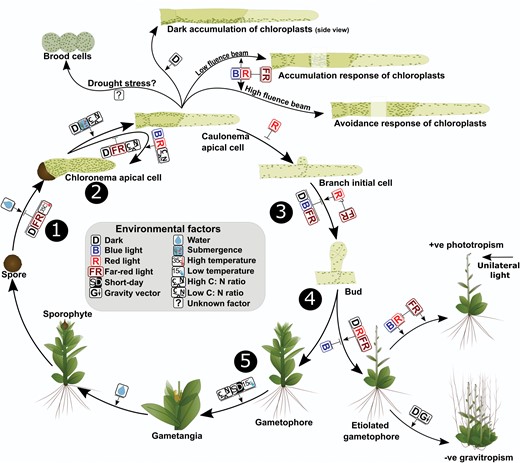
Effect of environmental signals on growth and development in mosses
academic.oup.com/jxb/article/73/13/4514/6565331
Mar 5, 2025
23
An Automation Workflow for High‐Throughput Manufacturing and Analysis of Scaffold‐Supported 3D Tissue Arrays - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC11468893/
Mar 5, 2025
7
BoneJ
imagej.net/plugins/bonej
Mar 1, 2025
2
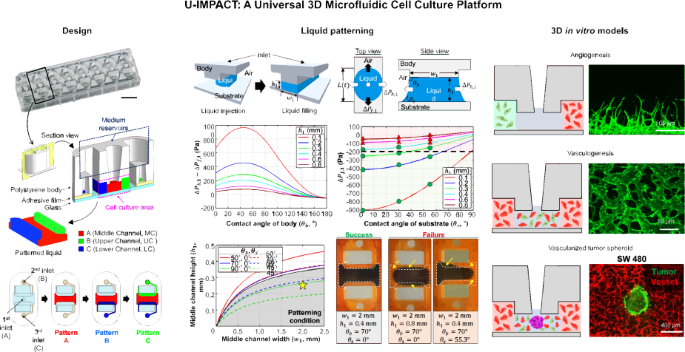
U-IMPACT: a universal 3D microfluidic cell culture platform - Microsystems & Nanoengineering
www.nature.com/articles/s41378-022-00431-w
Feb 28, 2025
2
Capillary-Driven Flow Microfluidics Combined with Smartphone Detection: An Emerging Tool for Point-of-Care Diagnostics - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC7459612/
Feb 28, 2025
5
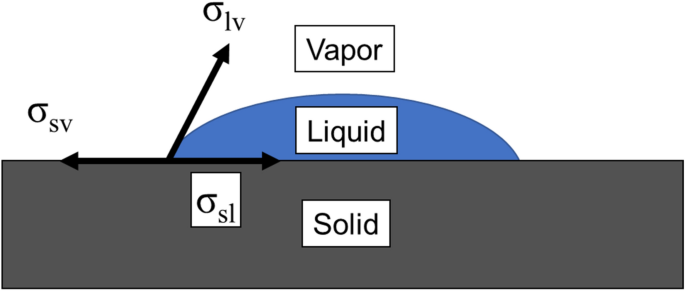
A general form of capillary rise equation in micro-grooves - Scientific Reports
www.nature.com/articles/s41598-020-76682-2
Feb 28, 2025
8
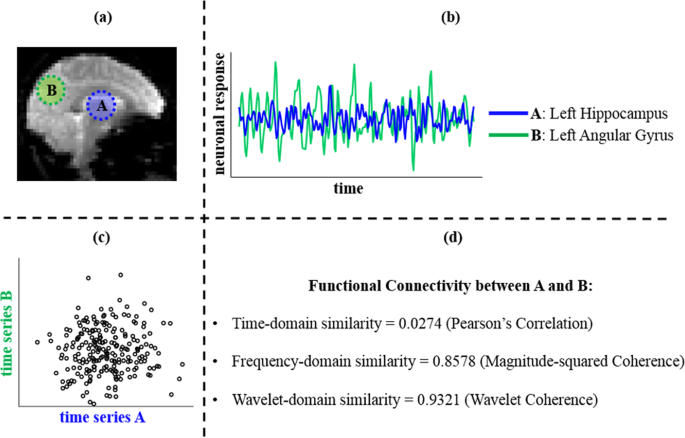
Rethinking Measures of Functional Connectivity via Feature Extraction - Scientific Reports
www.nature.com/articles/s41598-020-57915-w
Feb 26, 2025
3
Microfluidic assay for simultaneous culture of multiple cell types on surfaces or within hydrogels - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC4035049/
Feb 26, 2025
2

IBM SPSS Statistics 30.0.0
www.ibm.com/docs/en/spss-statistics/30.0.0?topic=data-statistics-table
Feb 13, 2025
1
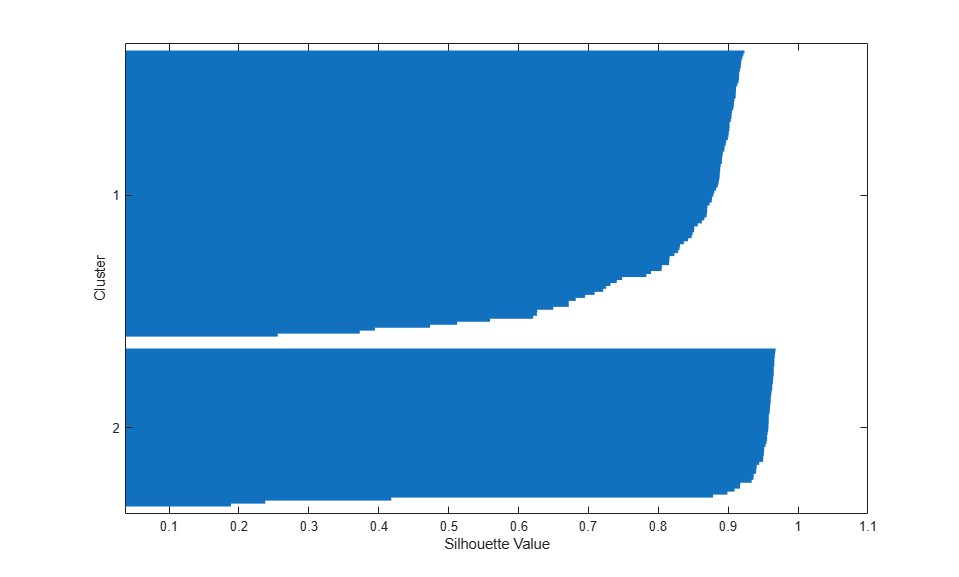
Cluster Analysis
www.mathworks.com/help/stats/cluster-analysis-example.html
Feb 13, 2025
7
Power-Law, Heavy & Fat-Tailed Distributions
rinterested.github.io/statistics/power_law.html
Feb 6, 2025
2

Making rose diagrams fit-for-purpose
www.sciencedirect.com/science/article/pii/S001282521930594X
Feb 5, 2025
2

Quantitative analysis of organelle distribution and dynamics in Physcomitrella patens protonemal cells - BMC Plant Biology
bmcplantbiol.biomedcentral.com/articles/10.1186/1471-2229-12-70
Feb 5, 2025
27

Immunofluorescence microscopy of microtubules in intact cell lineages of the moss, Physcomitrella patens: I. normal and cipc-treated tip cells
journals.biologists.com/jcs/article-split/75/1/131/59701/Immunofluorescence-microscopy-of-microtubules-in
Feb 5, 2025
12
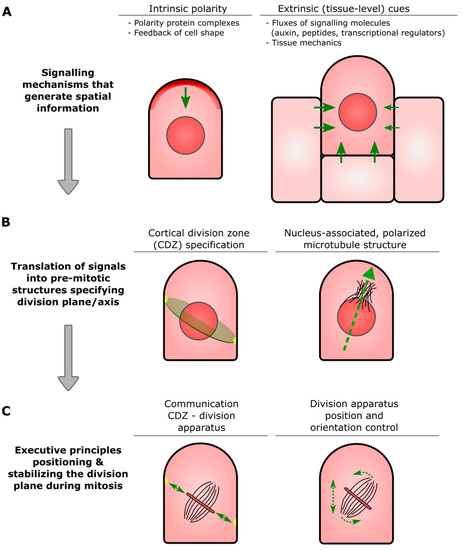
Physcomitrium patens: A Single Model to Study Oriented Cell Divisions in 1D to 3D Patterning
www.mdpi.com/1422-0067/22/5/2626
Feb 5, 2025
26
Symmetry Breaking in Plants: Molecular Mechanisms Regulating Asymmetric Cell Divisions in Arabidopsis - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC2773642/
Feb 5, 2025
2
Moss Pathogenesis-Related-10 Protein Enhances Resistance to Pythium irregulare in Physcomitrella patens and Arabidopsis thaliana - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC4850436/
Feb 5, 2025
2

Two new fluorescent dyes applicable for visualization of fungal cell walls
www.tandfonline.com/doi/epdf/10.1080/15572536.2006.11832788?needAccess=true
Feb 5, 2025
4

Application of Membrane and Cell Wall Selective Fluorescent Dyes for Live-Cell Imaging of Filamentous Fungi
app.jove.com/v/60613/application-membrane-cell-wall-selective-fluorescent-dyes-for-live
Feb 5, 2025
3

Concentration, cellular exposure and specificity of organelle selective fluorescent dyes in fungal cell biology
www.sciencedirect.com/science/article/pii/S1749461321000373
Feb 5, 2025
5

Real-Time Imaging of Cellulose Reorientation during Cell Wall Expansion in Arabidopsis Roots
academic.oup.com/plphys/article/152/2/787/6108732
Feb 5, 2025
9
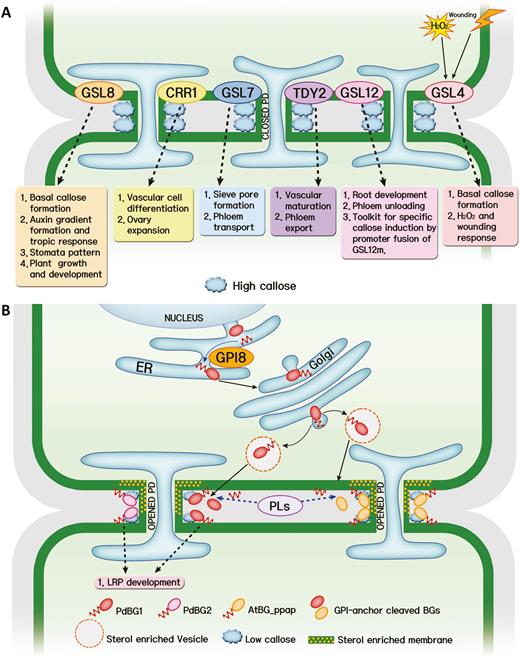
Callose balancing at plasmodesmata
academic.oup.com/jxb/article/69/22/5325/5085387
Feb 5, 2025
19

Callose balancing at plasmodesmata | Oxford Academic
academic.oup.com/view-large/figure/126192502/ery31702.jpg
Feb 5, 2025
1

CalTrack: High-Throughput Automated Calcium Transient Analysis in Cardiomyocytes | Circulation Research
www.ahajournals.org/doi/full/10.1161/CIRCRESAHA.121.318868
Feb 5, 2025
1
CalTrack: High-Throughput Automated Calcium Transient Analysis in Cardiomyocytes | Circulation Research
www.ahajournals.org/doi/suppl/10.1161/CIRCRESAHA.121.318868
Feb 5, 2025
1
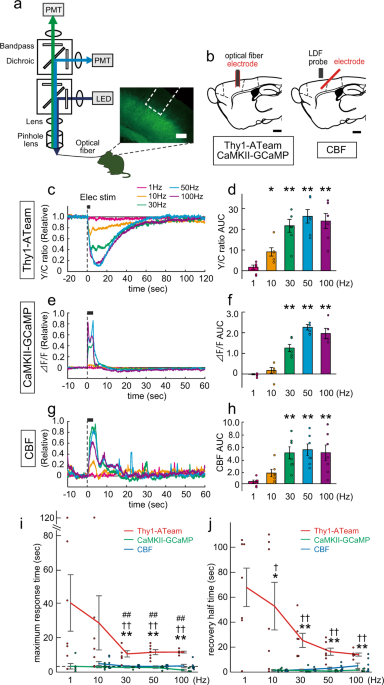
Intracellular ATP levels in mouse cortical excitatory neurons varies with sleep–wake states - Communications Biology
www.nature.com/articles/s42003-020-01215-6
Feb 5, 2025
3
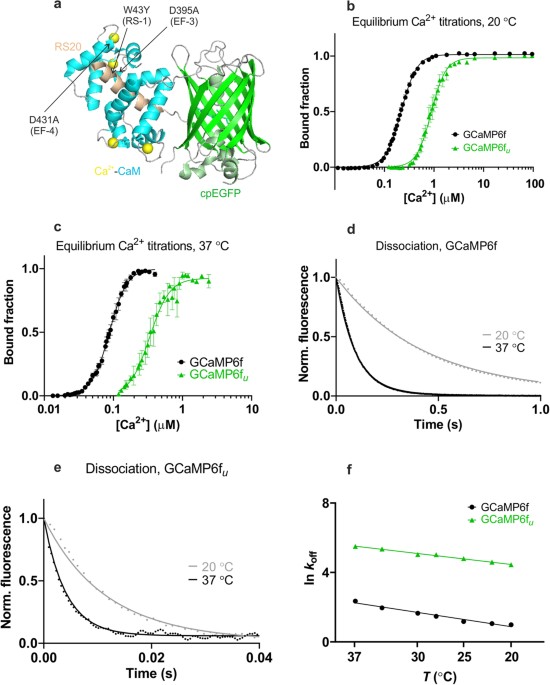
Design and mechanistic insight into ultrafast calcium indicators for monitoring intracellular calcium dynamics - Scientific Reports
www.nature.com/articles/srep38276
Feb 5, 2025
2
Conserved role of dopamine in the modulation of behavior - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC3502204/
Jan 31, 2025
19

Dopamine modulates the plasticity of mechanosensory responses in Caenorhabditis elegans | The EMBO Journal
www.embopress.org/doi/full/10.1038/sj.emboj.7600057
Jan 30, 2025
1

Noninvasive electrical stimulation as a neuroprotective strategy in retinal diseases: a systematic review of preclinical studies - Journal of Translational Medicine
translational-medicine.biomedcentral.com/articles/10.1186/s12967-023-04766-4
Jan 30, 2025
1
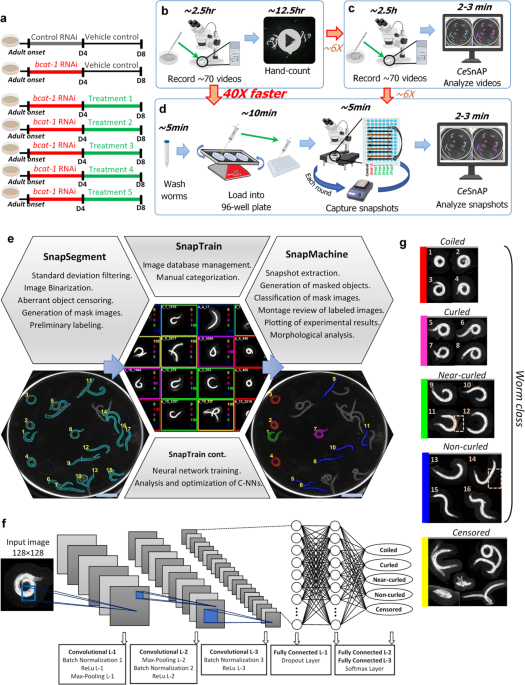
High-throughput behavioral screen in C. elegans reveals Parkinson’s disease drug candidates - Communications Biology
www.nature.com/articles/s42003-021-01731-z
Jan 30, 2025
3
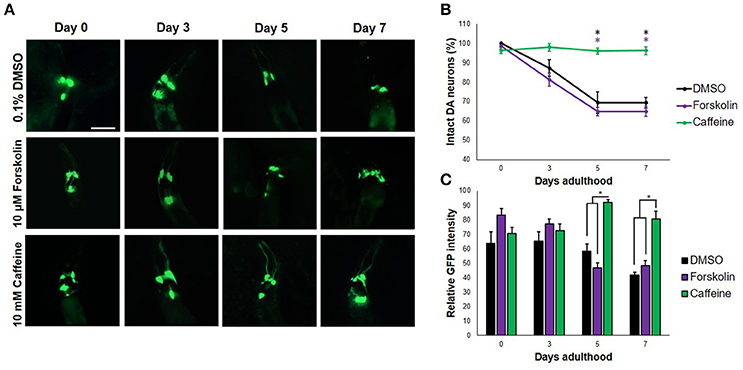
Frontiers | Caffeine Protects Dopaminergic Neurons From Dopamine-Induced Neurodegeneration via Synergistic Adenosine-Dopamine D2-Like Receptor Interactions in Transgenic Caenorhabditis elegans
www.frontiersin.org/journals/neuroscience/articles/10.3389/fnins.2018.00137/full
Jan 30, 2025
4
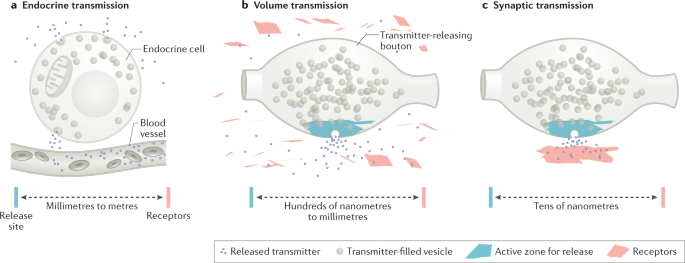
Spatial and temporal scales of dopamine transmission - Nature Reviews Neuroscience
www.nature.com/articles/s41583-021-00455-7
Jan 30, 2025
8
Extracellular Dopamine Potentiates Mn-Induced Oxidative Stress, Lifespan Reduction, and Dopaminergic Neurodegeneration in a BLI-3–Dependent Manner in Caenorhabditis elegans
journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1001084
Jan 30, 2025
1
A microfluidic phenotype analysis system reveals function of sensory and dopaminergic neuron signaling in C. elegans electrotactic swimming behavior - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC3704449/
Jan 30, 2025
354
Optogenetic manipulation of neural activity in freely moving Caenorhabditis elegans - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC3032981/
Jan 30, 2025
1
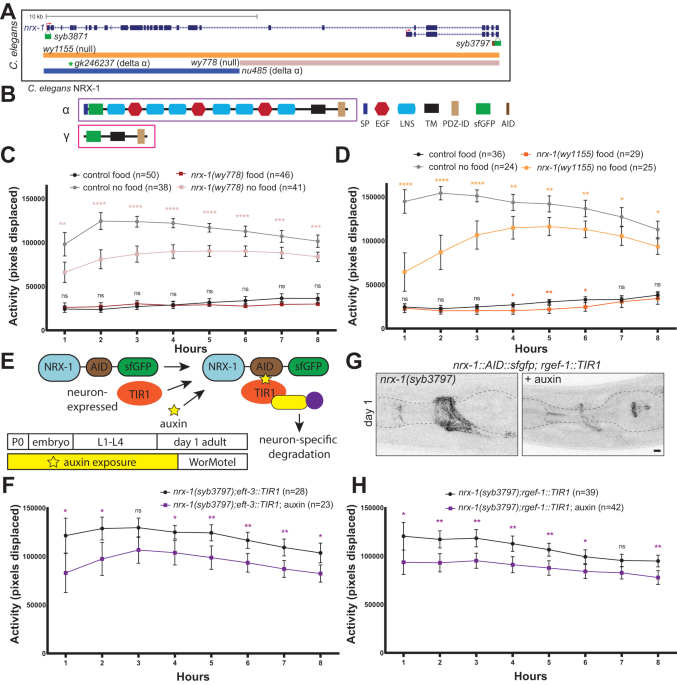
Distinct neurexin isoforms cooperate to initiate and maintain foraging activity - Translational Psychiatry
www.nature.com/articles/s41398-023-02668-z?fromPaywallRec=false
Jan 29, 2025
4
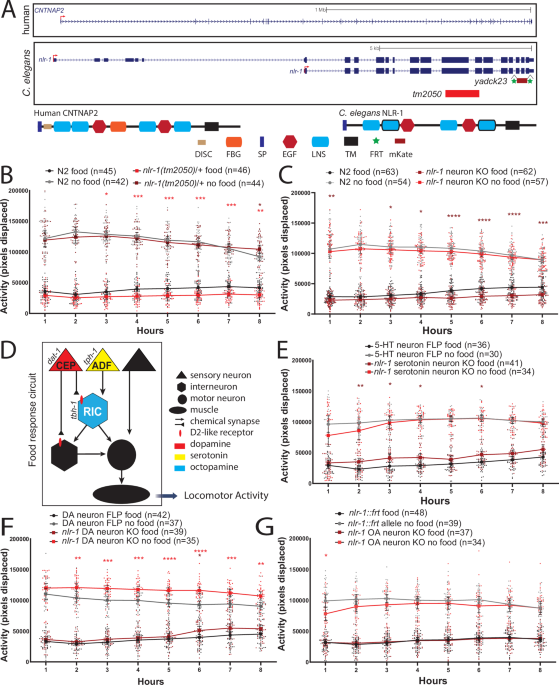
nlr-1/CNTNAP regulates dopamine circuit structure and foraging behaviors in C. elegans - Communications Biology
www.nature.com/articles/s42003-024-06936-6?fromPaywallRec=false
Jan 29, 2025
14
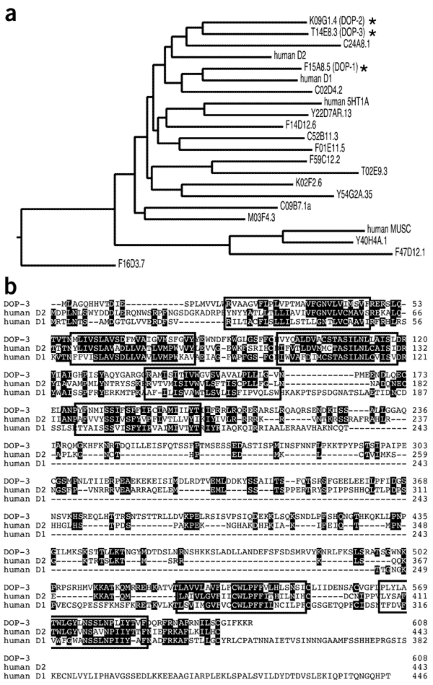
Mechanism of extrasynaptic dopamine signaling in Caenorhabditis elegans - Nature Neuroscience
www.nature.com/articles/nn1316
Jan 29, 2025
32
Stimulation of subterritories of the subthalamic nucleus reveals its role in the integration of the emotional and motor aspects of behavior - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC1965569/
Jan 29, 2025
1
The role of neurotransmitter systems in mediating deep brain stimulation effects in Parkinson’s disease - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC9579467/
Jan 29, 2025
4
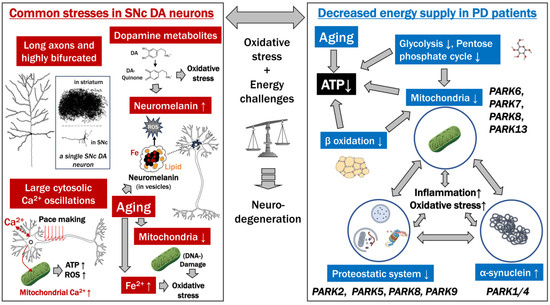
Parkinson’s Disease: Cells Succumbing to Lifelong Dopamine-Related Oxidative Stress and Other Bioenergetic Challenges
www.mdpi.com/1422-0067/25/4/2009
Jan 29, 2025
3
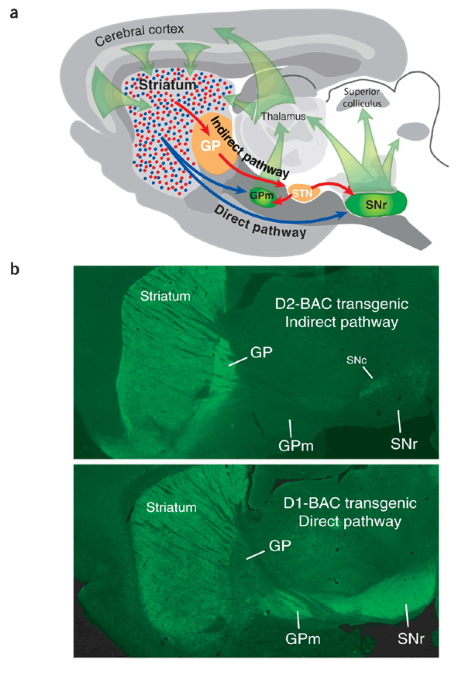
Indirect-pathway neurons lose their spines in Parkinson disease - Nature Neuroscience
www.nature.com/articles/nn0206-157
Jan 29, 2025
4
Myosin Storage Myopathy in C. elegans and Human Cultured Muscle Cells - PMC
pmc.ncbi.nlm.nih.gov/articles/PMC5268365/
Jan 29, 2025
3

Anesthetic-like effects of ketamine in C. elegans
www.sciencedirect.com/science/article/pii/S0306452224006304
Jan 29, 2025
10